Страница: 3/8
абсолютное; в нек-рых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответств. участками; этот процесс носит назв. перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетич. анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Генетических картах хромосом выражают как процент кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему кол-ву изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1 %.
Итак, выделим основные положения хромосомной теории наследственности:
1. Гены располагаются в хромосомах, различные хромосомы содержат неодинаковое чис ло генов, набор генов каждой из негомологичных хромосом уникален.
2. Гены в хромосоме расположены линейно, каждый ген занимает в хромосоме определенный локус (место).
3. Гены , расположенные в одной хромосоме, образуют группу сцепления и вместе (сцеплено) передаются потомкам, число групп сцепления равно гаплоидному набору хромосом.
4. Сцепление не абсолютно, так как в профазе мейоза может происходить кроссинговер и гены, находящиесяв одной хромосоме, разобщаются. Сила сцепления зависит от расстояния между генами в хромосоме: чем больше расстояние, тем меньше сила сцепления. и наоборот. Расстояние между генами измеряется в процентах кроссинговера. 1% кроссинговера соответтствует одной морганиде.[4]
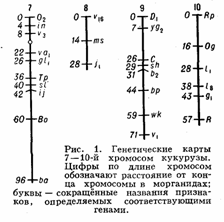
Генетические карты хромосом составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые назв. мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Генетические карты хромосом можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределённых в 10 группах сцепления (рис. 1). У менее изученных объектов число обнаруженных групп сцепления
меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом.
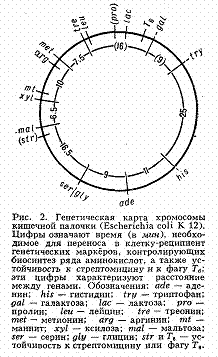
У бактерий, к-рые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетич. материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Генетических карт хромосом бактерий, детально разработанных у ряда видов. Ещё более детализированы Генетические карты хромосом нек-рых бактериофагов
Генетика пола. Количество групп сцепленных генов оказалось равным количеству пар хромосом, присущих данному виду. Важнейшие доказательства хромосомной теории наследственности были получены при изучении наследования, сцепленного с полом. Ранее цитологи открыли в хромосомных наборах ряда видов жиивотных особые , так называемые половые хромосомы, которыми самки отличаются от самцов. В одних случаях самки имеют 2 одинаковые половые хромосомы(XX), а самцы-разные(XY), в других - самцы-2 одинаковые(XX, или ZZ), а самки - разные(XY, или ZW). Пол с одинаковыми половыми хромосомами называется гомогаметным , с разными - гетерогаметным. Женский пол гомогаметен , а мужской гетерогаметен у некоторых насекомых ( в том числе у дрозофилы) и всех млекопитающих. Обратное соотношение - у птиц и бабочек. Ряд признаков у дрозофилы наследуется в
строгом соответствии с передачей потомству X-хромосом. Самка дрозофилы, проявляюща
рецесивный признак , например белую окраску глаз, в силу гомозиготности по этому гену, находящимуся в X-хромосоме, передает белую окраску глаз всем сыновьям, так как они получают свою X-хромосому только от матери. В случае гетерозиготности по рецессивному сцепленному с полом признаку самка передает его половине сыновей. При противоположном определении пола (самцы XX, или ZZ; самки-XY, или ZW) особи мужского пола передают сцепленные с полом признаки дочерям, получающим свою X( =Z ) хромосому от отца. Иногда в результате нерасхождения половых хромосом при мейозе возникают самки строения XXY и самцы XYY. Возможны также случаи соединения X-хромосом концами; тогда самки передают сцепленные X-хромосомы своим дочерям, у которых и проявляются сцепленные с полом признаки. Сыновья же похожи на отцов (такое наследование называется гологеническим ). Если наследуемые гены находятся в Y-хромосоме, то определяемые ими признаки передаются только по мужской линии - от отца к сыну (такое наследование называется голандрическим). Хромосомная теория наследственности вскрыла внутриклеточные механизмы наследственности, дала точное и единое объяснение всех явлений наследования при половом размножении, объяснила сущность изменений наследственности, то есть изменчивости.
Нехромосомная теория наследственности. Первенствующая роль ядра и хромосом в наследственности не исключает передачи некоторых признаков и через цитоплазму, в которой обнаружены структуры, способные к самовоспроизведению.Единицы цитоплазматической (нехромосомной) наследственности отличаются от хромосомных тем, что они не расходятся при мейозе. Поэтому потомство при нехромосомной наследственности воспроизводит признаки только одного из родителей (чаще матери ). Таким образом , различают ядерную наследственность, связанную с передачей наследственных признаков, находящихся в хромосомах ядра (иногда ее называют хромосомной наследственностью ), и внеядерную наследственность, зависящую от передачи самовоспроизводящихся структур цитоплазмы. Ядерная наследственность реализуется и при вегетативном размножении , но не сопровождается перераспределением генов, что наблюдается при половом размножении, а обеспечивает константную передачу признаков из поколения в поколение, нарушаемую только соматическими мутациями.
Молекулярная генетика. Применение новых физических и химических методов, а также использование в качестве объектов исследования бактерий и вирусов резко повысили разрешающую способность генетических экспериментов, привели к изучению наследственности на молекулярном уровне и бурному развитию молекулярной генетики. Впервые Н. К. Кольцов (1927 г) выдвинул и обосновал представления о молекулярной основе наследственности и о матричном способе размножения “наследственных молекул”.В 40-х гг. 20 в. была экспериментально доказана генетическая роль дизоксирибонуклеиновой кислотиы ( ДНК ) , а в 50-60-х гг. установлена ее молекулярная структура и выяснены принципы кодирования генетической информации. Генетическая информация,заложенная в наследственных структурах организмов (в хромосомах, цитоплазме, клеточных организмах), получаемая от предков в виде совокупности генов информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот) и связанных с ними функциях. У многоклеточных форм при половом размножении генетическая информация передаётся из поколения в поколение через посредство половых клеток — гамет, единственная функция к-рых — передача и хранение генетической информации. У микроорганизмов и вирусов имеются особые типы ее передачи . Генетическая информация заключена преимущественно в хромосомах, где она зашифрована в определённой линейной последовательности нуклеотидов в молекулах дезоксирибонуклеиновой кислоты — ДНК (генетический код). Генетический код - это система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах — дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК)—встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой к-те), разлчающихся по входящему в их состав азотистому основанию . В ДНК встречаются основания:
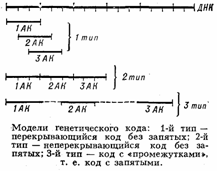
аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых к-т обнаружено ок. 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных Сахаров. Так как количество кодирующих знаков Генетического кода (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. кол-во нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42 = 16, но этого также недостаточно для зашифровки всех аминокислот. Американский учёный Г. Гамов предложил (1954) модель т р и п л е т н о г о генетического кода, т. е. такого, в котором 1 аминокислоту кодирует группа из трёх нуклеотидов, называемых кодоном. Число возможных триплетов равно 43 = 64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует несколько кодонов (так называемая вырожденность кода). Было предложено много различных моделей генетического кода, из которых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены след. осн. закономерности, касающиеся генетического кода: 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (коллинеарность генетического кода); 2) считывание кода начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) генетический код, как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность генетического кода уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём;
Реферат опубликован: 23/05/2005 (18628 прочтено)