Страница: 4/4
Построение калибровочной кривой исключает ошибки, происходящие от качественных различий реакции аминокислот на цветной реактив. До оптической плотности 0.1 зависимость между величиной поглощения и количеством субстрата носит линейный характер.
Воспроизводимость составляет ±3%. Мешающее влияние аммония, аминов и других веществ, содержащихся в биологическом материале и дающих цветную реакцию с нингидридом, исключается при помощи соответствующих холостых проб. Анализ сравнительно упрощается, когда изучают протеолитические ферменты, субстратами которых являются пептиды, не содержащие свободных аминогрупп. В этом случае вся образующаяся окраска целиком приходится на освободившуюся аминокислоту.
Микрометод определения активности протеиназ А.П.Алексеенко. Предлагаемый микрометод определения активности протеолитических ферментов основан на измерении содержания аргинина в пептидах, освобождающихся при гидролизе протеиназами белковых субстратов. Содержание аргинина определяют с помощью стабилизированной и усовершенствованной реакции Сакагучи (хроматограмму погружают в 0.1% раствор 8-гидроксихинолина в ацетоне, высушивают на воздухе, опрыскивают раствором 0.2 мл Br2 в 100 мл 0.5 М NaOH; аргинин и другие гуанидины дают оранжево-красные пятна, тауромицин и гликоциамин - лишь временную окраску), позволяющей определить содержание аргинина в растворах трихлоруксусной кислоты после осаждения и удаления нерастворимых белков. Зная содержание аргинина в белковом субстрате и определяя его количество в перешедших в раствор пептидах, можно рассчитать процент гидролиза белкового субстрата изучаемыми протеиназами. В этом состоит главное преимущество метода, поскольку метод Ансона не дает возможности рассчитать процент гидролиза белка, атакуемого протеолитическими ферментами. Из всех существующих методов только метод Мура и Штейна позволяет определить процент гидролиза белковых субстратов, но он имеет ограниченное применение, поскольку требует предварительного проведения полного кислотного гидролиза как белка-субстрата, так и образовавшихся пептидов. В предложенной методике калибровочная кривая строится по растворам свободного аргинина. Введение в реакцию пептидов и белков в виде их биуретовых комплексов повысило чувствительность реакции Сакагучи с остатками аргинина и сделало ее такой же чувствительной, как и для свободного аргинина. В качестве белкового субстрата используют классический субстрат - гемоглобин и окисленный по Сангеру лизоцим. Этот низкомолекулярный белок, содержащий 12.7% аргинина прекрасно расщепляется пепсином (рвется около 30 связей).
Прежде чем приступить к определению активности ферментов в изучаемых тканевых субстратах, необходимо:
1. Определить зависимость активности от рН и выбрать оптимальную реакцию среды для проявления активности изучаемого фермента.
2. Построить график зависимости активности фермента от времени его инкубации с субстратом. выбрать для работы те сроки инкубации, при которых сохраняется линейная зависимость между величиной активности фермента и временем его инкубации.
3. Определить зависимость величины активности от концентрации белка в пробе. Выбрать такие концентрации фермента, при которых величина активности фермента была бы пропорциональна его концентрации. Следует учитывать, что в тканевых экстрактах и биологических жидкостях могут присутствовать ингибиторы протеолитических ферментов. При разведении пробы их концентрация снижается , а протеолитических - увеличивается. Определяя “фактор разведения”, можно одновременно с подбором оптимальных условий для определения протеолитической активности выявить и наличие ингибитора.
4. При определении активности фермента необходимо работать при постоянной скорости ферментативной реакции, достигаемой при полном насыщении фермента субстратом, при так называемой максимальной скорости ферментативной реакции. В каждом отдельном случае максимальную скорость необходимо найти экспериментально, измерив активность препарата при разных концентрациях субстрата, при постоянных концентрациях белка и времени инкубирования.
Определение активности протеиназ по белковому субстрату.
К 0.5 мл субстрата в соответствующем буферном растворе добавляют 0.5 мл пробы, содержащей фермент (экстракт, биологическую жидкость), инкубируют установленное опытным путем время , после чего белки в пробе осаждают 5 мл 10% трихлоруксусной кислотой. Отделяют осадок центрифугированием, а надосадочную жидкость (ТХУ-центрифугат) подвергают дальнейшей обработке. Контрольные пробы - пробы, где реактив добавлен в обратном порядке: к 3 мл 10% раствора ТХУ добавляют 0.5 мл раствора, содержащего фермент и 0.5 мл субстрата. Рекомендуется ставить пробу на автолиз субстрата: к 0.5 мл субстрата добавляют 0.5 мл прокипяченного раствора, содержащего фермент, инкубируют вместе с опытными пробами и осажденной ТХУ.
Определение содержания аргинина в ТХУ-центрифугантах по модифицированной реакции Сакагучи. К 0.5 мл ТХУ-центрифуганта добавляют 0.5 мл 2.5мМ раствора CuSO4 , 0.5 мл 5 Н. КОН, 0.5 мл раствора ДХН (2,4-дихлор-1-нафтол). Раствор встряхивают и вносят 0.5 мл гипобромита натрия, мгновенно возникает ярко-розовая окраска. Раствор встряхивают и через 20-30 секунд стабилизируют пробы добавлением 0.2 мл раствора 2-тиогликоля (чтобы предотвратить разрушение окрашенного продукта избытком гипобромита натрия. После добавления 2-тиодигликоля в пробы как опытные, так и контрольные, появляется желтоватое окрашивание за счет побочного продукта реакции между избыточной ТХУ и гипобромита натрия. Побочный продукт также стабилизируется 2-тиодигликолем. Оптическая плотность пробы измеряется спектрофотометром при 520 нм в кювете толщиной 1 см.
Определение активности протеиназ по реакции с реактивом Фолина. К 0.5 мл того же ТХУ-центрифуганта добавляют 0.5 мл 2.5 мМ раствора CuSO4 , 4 мл 0.5 н. раствора NaOH и 1.5 мл разбавленного в три раза реактива Фолина. Через 30 минут измеряют на спектрофотометре оптическую плотность при 760 нм
Калибровочные кривые по пепсиновому гидролизату окисленного лизоцима. 30 мг окисленного лизоцима растворяют в 50 мл подкисленной воды (40 мл Н2О + 10 мл 0.3 н. раствора HСl. и к полученном раствору добавляют 0.5 мг пепсина. Смесь оставляют при комнатной температуре на сутки. По истечении этого срока инкубации препарат не дает осадка с ТХУ. Этот препарат используют наряду с растворами аргинина и тирозина в качестве стандарта для построения калибровочных кривых. Приготовляют разведения с содержанием от 60 до 600 мкг/мл исходного лизоцима. В пробы вводят соответствующее опытным пробам количество ТХУ. Приготовляют также растворы свободного аргинина и тирозина с концентрациями от 0.04 до 0.25 мкмоль/мл. Из каждой пробы берут по 0.5 мл (в трех параллельных пробах) для определения содержания аргинина и тирозина по Фолину.
Калибровочные кривые. 3 получена с помощью микрометода на основе реакции Сакагучи (520 нм) по растворам аргинина (х) и по пептидному гидролизу лизоцима (о); 1 и 2 - с помощью реакции Фолина (760 нм): 2 - по растворам тирозина, 1 - по пептидному гидролизу лизоцима. Значение оптической плотности
проб, измеренные при 520 нм или 760 нм, нанесены на график против концентрации
аргинина или тирозина или их остатков в пепсиновом гидролизате лизоцима. Кривая
1, проходящая выше кривой 2, получена при реакции реактива Фолина с рас-
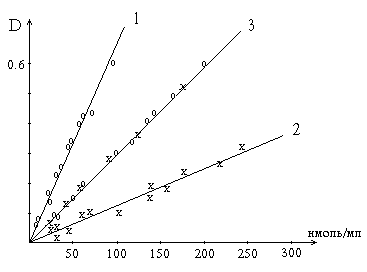
Количество аминокислоты или ее остатка
в гидролизате в наномолях на 1 мл.
творами тирозина эквивалентной концентрации. Кривая 3 получена в результате реакции Сакагучи как со свободным аргинином, так и с его остатками, содержащимися в пептидах пепсинового гидролизата лизоцима. Расположение опытных точек точно по прямой свидетельствует о том, что в гидролизате, осажденном ТХУ, весь аргинин полностью реагирует с реактивом Сакагучи.
Воздействие Фолина на тирозин не специфично. Кроме тирозина в пептидах с этим реактивом реагируют триптофан, цистеин. Образуемые медью с тетрапептидами и полипептидами биуретовые комплексы облегчают такое взаимодействие. Следовательно, выражение величины активности протеолитических ферментов в тирозиновых эквивалентах и по Фолину, как это иногда делают, неправильно.
Таким образом, мы рассмотрели особенности ферментов как биологических катализаторов, показаны их отличия от небелковых катализаторов, способы измерения активности предложенных ферментов - пепсина и папаина. К сожалению, на данный момент имеется довольно незначительное количество публикаций по исследованию папаина.
Литература
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник.- М.: Медицина, 1990.- с.115
2. Основы биохимии: Учебник для студ. биол. спец. ун-тов/под ред. А.А. Анисимова.- М.: Выс.шк., 1986. - с.133-140
3. Фёршт Э. Структура и механизм действия ферментов.- М.: Мир., 1980.- с. 373-388
4. Кочетов Г.А. Практическое руководство по энзимологии.- М., 1989
5. Диксон М., Уэбб Э. Ферменты: пер.англ.- М.: Мир, 1982.- т.1.- с. 370-375
6. Асатиани В.С. Биохимическая фотометрия.- М.: Изд. АН СССР, 1957.- с.248-253
Реферат опубликован: 1/06/2005 (13515 прочтено)