Страница: 4/7
Ф. Арриги и соавторы (1971) установили, что уникальные последовательности занимают более 56% ДНК хромосом человека, высокоповторяющиеся — 12,4 %, промежуточные повторы — 8 %. Общее количество повторяющихся генов в ДНК хромосомы человека равно 28%. Число хромосом у человека длительное время оставалось невыясненным. Дело в том, что определить количество хромосом у млекопитающих, особенно у человека, было трудно. Хромосомы оказались маленькими, весьма многочисленными, плохо поддавались подсчету. При фиксации клетки они сливались в комки, что затрудняло определение истинного числа хромосом. Поэтому первые исследователи не могли точно и правильно подсчитать количество хромосом в клетках человека. Называлось разное количество хромосом — от 44 до 50.
![]()
 Обычно
хромосомы в клетках наблюдают во время митоза на стадии метафазной пластинки.
В интерфазном ядре хромосомы в световой микроскоп не видны. В 1912 г. Г. Винивартер,
изучая хромосомы в сперматогониях и оогониях половых желез человека, удаленных
во время операции, установил, что мужской набор хромосом (кариотип) содержит
47 хромосом, а женский — 48. В 1922 г. Т. Пайнтер повторил исследования Винивартера
и установил, что мужской и женский кариотипы содержат по 48 хромосом, но женский
отличается от мужского только двумя хромосомами. У женщин находится 2 большие
половые хромосомы, а у мужчины одна большая Х-хромосома и одна маленькая К-хромосома.
В последующие годы эту точку зрения поддерживали и другие ученые. П. И. Живаго
и А. Г. Андреа (1932) предложили первую классификацию хромосом в зависимости
от их длины. Так как хромосомы очень близко располагаются одна около другой
и их очень трудно исследовать, то и в последующие годы точное число хромосом
у человека служило предметом споров и дискуссий. Однако постепенно было достигнуто
согласие между исследователями по этому вопросу, и в течение 30 лет большинство
цитогенетиков считало, что у человека диплоидное число хромосом равно 48, а
гаплоидное — 24. Усовершенствованные методы изучения хромосом позволили получить
более точные сведения о количестве хромосом в клетках у человека, а также выявить
аномалии нормального кариотипа, ответственные за некоторые уродства. Особенно
плодотворным оказались два метода:
Обычно
хромосомы в клетках наблюдают во время митоза на стадии метафазной пластинки.
В интерфазном ядре хромосомы в световой микроскоп не видны. В 1912 г. Г. Винивартер,
изучая хромосомы в сперматогониях и оогониях половых желез человека, удаленных
во время операции, установил, что мужской набор хромосом (кариотип) содержит
47 хромосом, а женский — 48. В 1922 г. Т. Пайнтер повторил исследования Винивартера
и установил, что мужской и женский кариотипы содержат по 48 хромосом, но женский
отличается от мужского только двумя хромосомами. У женщин находится 2 большие
половые хромосомы, а у мужчины одна большая Х-хромосома и одна маленькая К-хромосома.
В последующие годы эту точку зрения поддерживали и другие ученые. П. И. Живаго
и А. Г. Андреа (1932) предложили первую классификацию хромосом в зависимости
от их длины. Так как хромосомы очень близко располагаются одна около другой
и их очень трудно исследовать, то и в последующие годы точное число хромосом
у человека служило предметом споров и дискуссий. Однако постепенно было достигнуто
согласие между исследователями по этому вопросу, и в течение 30 лет большинство
цитогенетиков считало, что у человека диплоидное число хромосом равно 48, а
гаплоидное — 24. Усовершенствованные методы изучения хромосом позволили получить
более точные сведения о количестве хромосом в клетках у человека, а также выявить
аномалии нормального кариотипа, ответственные за некоторые уродства. Особенно
плодотворным оказались два метода:
1. Обработка культуры клеток алкалоидом колхицином, который ведет к накоплению делящихся клеток на стадии метафазы;
2. Обработка клеток слабыми растворами солей, вызывающими набухание, расправление хромосом, что облегчает их исследование.
В 1956 г. шведские цитологи Дж. Тийо и А. Леван изготовили культуры клеток из тканей легких, взятых у абортированных человеческих эмбрионов и, используя усовершенствованную методику обработки клеток, получили необычайно четкие препараты, в которых ясно было видно 46 хромосом.[5]
Несколькими месяцами позднее Ч. Форд и Дж. Хаммертон в Англии установили, что диплоидные предшественники половых клеток в семенниках мужчин (сперматогонии) также имеют по 46 хромосом, а гаплоидные (сперматоциты 1-го деления) — по 23 хромосомы.
После этого были изучены многие клетки из разных органов и тканей человека и везде нормальное число хромосом оказалось равным 46.
Женский кариотип отличается от мужского только одной половой хромосомой. Остальные 22 пары одинаковы у мужчин и женщин. Эти 22 пары хромосом называются аутосомами. Нормальный кариотип состоит из 44 аутосом (22 пары) и двух половых хромосом — XX у женщин и XY у мужчин, т. е. женский кариотип имеет две большие половые хромосомы, а мужской — одну большую и одну маленькую.
В половых клетках человека находится одинарный (гаплоидный) набор хромосом — 23, а в соматических клетках — двойной (диплоидный) набор — 46. Эти открытия стимулировали дальнейшее изучение хромосом. Были разработаны методы исследования хромосом в культуре лимфоцитов периферической крови и на других объектах. В настоящее время хромосомы относительно легко исследуют в лимфоцитах периферической крови. Венозную кровь помещают в специальную питательную среду, добавляют фитогемаглютинин, который стимулирует клетки к делению, и помещают на 72 ч. в термостат. За 6 ч. до конца инкубации сюда добавляют колхицин, который задерживает процесс деления клеток на стадии метафазной пластинки. Затем культуру помещают в гипотонический раствор NaCl, в котором клетки набухают, что приводит к легкому разрыву оболочек ядра и переходу хромосом в цитоплазму. После этого препараты окрашивают ядерными красителями, в частности ацетоорсеином, и рассматривают их в световом микроскопе с иммерсией.
Под микроскопом учитывают общее количество хромосом, фотографируют их, затем из фото вырезают ножницами каждую хромосому и наклеивают на чистый лист бумаги в ряд, начиная от самой большой (первой) хромосомы и кончая самой маленькой (двадцать второй) и половой Y-хромосомой. Люминесцентная методика позволяет быстро и просто проводить массовые исследования с целью выявления больных с различными типами хромосомных аномалий. Совокупность количественных (число хромосом и их размеры) и качественных (морфология хромосом) признаков диплоидного набора единичной клетки обозначается термином «кариотип». Строение хромосом изменяется в зависимости от стадии деления клеток (профазы, метафазы, анафазы, телофазы).
Уже в профазе митоза видно, что хромосома образована двумя взаимно переплетающимися нитями одинакового диаметра — хроматидами. В метафазе хромосома уже спирализована, и две ее хроматиды ложатся параллельно, разделенные узкой щелью. Каждая хроматида состоит из двух полухроматид. В результате митоза хроматиды материнской хромосомы становятся сестринскими хромосомами, а полухроматиды — их хроматидами. В основе хроматид лежат хромонемы — так называют более тонкие нити ДНП, состоящие из белка и нуклеиновых кислот.
В интерфазе (промежуток между двумя делениями клеток) хроматин тесно связан
с ядерными мембранами и ядерным белковым матриксом. Он образует также большие
участки деспирализованных нитей ДНП. Затем постепенно хроматин спирализуется,
образуя типичные метафазные
 хромосомы.
Размеры их варьируют от 2 до 10 микрон.
хромосомы.
Размеры их варьируют от 2 до 10 микрон.
В настоящее время интенсивно исследуются структурные особенности аутосом и половых хромосом (на клетках костного мозга, лимфоцитах, фибробластах, клетках кожи, регенерирующей печени).
![]() В хромосомах
выявлены структуры, названные хромомерами. Хромомер — это спирализованный участок
хромонемы. Промежутки между хромомерами представлены хромонемными нитями. Расположение
хромомеров на каждой хромосоме строго фиксировано, наследственно детерминировано.
В хромосомах
выявлены структуры, названные хромомерами. Хромомер — это спирализованный участок
хромонемы. Промежутки между хромомерами представлены хромонемными нитями. Расположение
хромомеров на каждой хромосоме строго фиксировано, наследственно детерминировано.
Хромомер — сравнительно крупная генетическая единица, сравнимая по длине с хромосомой кишечной палочки. Строение и функция хромомера — основная загадка современной генетики. Предполагают, что некоторые хромомеры — это один генетический локус, где есть один структурный ген и много генов регуляторных. Возможно, в других хромомерах располагается несколько структурных генов.
Хромонемы и хромомеры окружены неокрашивающимся веществом — матриксом. Полагают, что матрикс содержит дезоксирибонуклеиновую и рибонуклеиновую кислоты, белки.
Определенные участки хромосом образуют ядрышки. Ядрышки — это более или менее деспирализованные участки хромосом, окруженные продуктами деятельности генов (рибосомы, частицы РНК и т. п.). Здесь идет синтез рибосомальной РНК, а также осуществляются определенные этапы формирования рибосом. В нем синтезируется большая часть РНК клетки.
В метафазной хромосоме различают еще несколько образований: центромеру, два плеча хромосомы, теломеры и спутник.
Центромерный (meros — по-гречески, часть) участок хромосомы — это неокрашивающийся разрыв в хромосоме, видимый на препарате хромосом. Центромера содержит 2—3 пары хромомер, имеет сложное строение. Предполагают, что она направляет движение хромосомы в митозе. К центромерам прикрепляются нити веретена.
Теломеры — специальные структуры на концах хромосом — также имеют сложное строение. В их состав входит несколько хромомер. Теломеры предотвращают концевое присоединение метафазных хромосом друг к другу. Отсутствие теломеров делает хромосому «липкой» — она легко присоединяется к другим фрагментам хромосом.
Одни участки хромосомы называются эухроматиновыми, другие — гетерохроматиновыми. Эухроматиновые районы хромосом — это генетически активные участки, они содержат основной комплекс функционирующих генов ядер. Потеря даже мельчайшего фрагмента эухроматина может вызвать гибель организма. Гетерохроматиновые районы хромосом — обычно сильно спирализованы и, как правило, генетически мало активны. В гетерохроматине находится ядрышковый организатор. Потеря даже значительной части гетерохроматина часто не приводит организм к гибели. Гетерохроматиновые участки хромосомы реплицируются позднее, чем эухроматиновые. Следует помнить, что эухроматин и гетерохроматин — это не вещество, а функциональное состояние хромосомы.
Если расположить фотографии гомологичных хромосом по мере возрастания их размеров, то можно получить так называемую идиограмму кариотипа. Таким образом, идиограмма — это графическое изображение хромосом. На идиограмме пары гомологов располагаются рядами в порядке убывающего размера.
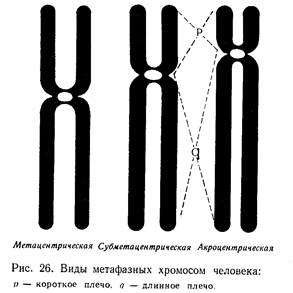
У человека на идиограмме среди 46 хромосом различают три типа хромосом в зависимости от положения в хромосоме центромер:
1. Метацентрические — центромера занимает центральное положение в хромосоме, оба плеча хромосомы имеют почти одинаковую длину;
2. Субметацентрические — центромера располагается ближе к одному концу хромосомы, в результате чего плечи хромосомы разной длины.
|
Классификация хромосом человека по размеру и расположению центромера |
||
|
Группа хромосом |
Номер по кариотипу |
Характеристика хромосом |
|
А(1) |
1,2,3 |
1 и 3 почти метацентрические и 2—крупная субметацентрическая |
|
В (11) |
4,5 |
крупные субакроцентрические |
|
С (III) |
6—12 |
средние субметацентрические |
|
A(lV) |
13—15 |
средние акроцентрические |
|
E(V) |
16-18 |
мелкие субметацентрические |
|
F(VI) |
19—20 |
самые мелкие мегацентрические |
|
G(VII) |
21—22 |
самые мелкие акроцентрические |
|
Х-хромосома (относится к III группе |
23 |
средняя почти метацентрическая |
|
Y-хромосома |
23 |
мелкая акроцентрическая |
Реферат опубликован: 15/04/2005 (38920 прочтено)