Страница: 3/4
Другие методы. Сюда относится обширный ряд методов, включающих поляриметрию, вискозиметрию, потенцио- и кондуктометрические измерения и т.п. Также определение активности можно выполнять, используя методы хроматографии и электрофореза на бумаге. Эти методы высокочувствительны и специфичны, что делает их во многих случаях незаменимыми; они позволяют значительно сократить расход фермента на измерение активности, но не всегда применимы ввиду продолжительности разделения веществ в процессе хроматографии (и электрофореза).
Специальные методы определения активности пепсина и папаина. Пепсин и папаин относятся к одной из наиболее многочисленных групп ферментов - протеолитические ферменты (протеазы). Они принадлежат к классу гидролаз, к подклассу пептидгидролаз и катализируют расщепление белков и полипептидов в соответствии с уравнением :
R1CONHR2 + H2O ® R1COOH + H2NR2 ,
R1 и R2 - остатки аминокислот, ди- или полипептиды.
Т.е. протеазы катализируют расщепление пептидной связи ¾CO¾NH¾ .
Теперь немного подробнее о пептидгидролазах.
Подкласс пептидгидролаз делится на следующие группы:
1. аминопептидазы (a-аминоацилпептидгидролазы) катализируют гидролиз полипептидов по месту пептидной связи, находящейся рядом со свободной аминной группой.
действие
R1 фермента R2
| ¯ |
H2N ¾ C ¾ C ¾ NH ¾ C ¾ C ¾
| ║ | ║
H O H O
2. карбоксипептидазы (пептидиламинокислотные гидролазы) катализируют расщепление полипептидов по месту пептидной связи, находящейся рядом со свободной гидроксильной группой.
R
¯ |
¾CO¾NH¾C¾H
|
действие COOH
фермента
3. дипептидазы (дипептидгидролазы) катализируют гидролитическое расщепление на свободные аминокислоты.
NH2CH2CONH¾CH2¾COOH + H2O ® 2CH2NH2COOH
глицил-глицин гликолол
Дипептидазы расщепляют только такие пептидные связи, по соседству с которыми находятся одновременно свободные карбоксильная и аминная группы. Большое влияние на действие дипептидаз оказывает наличие a-водородных атомов, находящихся у атомов углерода, связанных со свободной карбоксильной и аминной группами. При замене этих атомов другими группировками активность ферментов снижается или совсем исчезает.
4. протеиназы - (пептидилгидролазы) гидролизуют непосредственно белок. При этом образуются полипептиды и свободные аминокислоты. К этой подгруппе протеиназ принадлежит пепсин, трипсин, химотрипсин, папаин и др. В процессе действия ферментов на субстраты решающую роль играет структура молекул субстрата (соответствующая часть молекулы субстрата), в которой происходит разрыв. Характер распада белковой молекулы на пептиды и аминокислоты зависит от природы субстрата и внешних условий (рН, температуры и др.). В качестве белковых субстратов используют желатин, гемоглобин, казеин, сывороточный альбумин.
Методы определения активности протеолитических ферментов
Модификационный метод. Модификационный метод с применением субстрата казеина основан на определении скорости ферментативной реакции гидролиза субстрата под действием исследуемых протеолитических ферментов, содержащихся в материале, взятом на анализ. Скорость реакции определяют по количеству образовавшихся аминокислот - тирозина (a-амино-b-оксифенилпропионовая кислота) и триптофана (a-амино-b-индолилпропионовая кислота), которые устанавливают колориметрической реакцией с реактивом Фолина. Этим методом определяют указанные аминокислоты как в свободном, так и в связанном состоянии.
По количеству тирозина и триптофана, содержащемуся в гидролизате, определяют количество превращенного белка в процессе ферментативной реакции (из расчета содержания в белке 6% тирозина и 1.8% триптофана). В основу расчета активности протеолитических ферментов положена зависимость степени гидролиза белка от числа единиц активности фермента, введенных в ферментативную реакцию. На основе этой зависимости составляется график.
Поскольку в данном методе количество аминокислот, характеризующих количество превращаемого белка, определяется по интенсивности окраски реакционных сред - оптической плотности после реакции с реактивом, то в графике для простоты расчета вместо количества превращенного белка ставится пропорциональная ему оптическая плотность раствора. При составлении графика по оси абсцисс откладывают число единиц фермента, введенное в ферментативную реакцию, а по оси ординат - оптическую плотность, полученную после колориметрической реакции с реактивом Фолина.
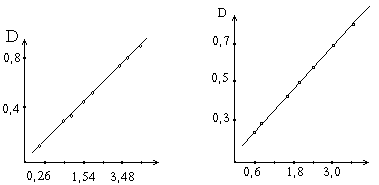
Содержание белка,мг Единицы активности
Зависимость оптической плотности от количества превращенного в процессе ферментативной реакции белка.
Зависимость оптической плотности от числа единиц активности протеолитических ферментов.
Относя количество введенных в реакцию единиц активности фермента к действию 1 г препарата, определяют его активность. Для расчета прямой, изображенной на графике, составляет расчетное уравнение.
За единицу протеолитической активности принято такое количество фермента, которое катализирует за 30 минут гидролиз 1 г белка в принятых условиях до продуктов, не осаждающихся трихлоруксусной кислотой. 1г составляет 25% от взятого на ферментативную реакцию белка.
Протеолитическая активность характеризуется числом единиц активности фермента, содержащегося в 1 г препарата и твердых полупродуктов или в 100 мл жидких материалов. Метод предназначен для анализа очищенных ферментных препаратов грибного происхождения и всех полупродуктов, получающихся при их производстве.
Определение пепсина по Ансону и Мирскому. Данный метод основан на следующих принципах. Гемоглобин подвергают воздействию пепсина, оставшийся гемоглобин осаждают трихлоруксусной кислотой. Фенольные свойства остатка (тирозин), определяемые фотометрически, используют как меру активности фермента.
Субстратом служит 2%-ный раствор гемоглобина в 0.06 н. растворе соляной кислоты, 5 мл субстрата нагревают до 250С, прибавляют 1 мл раствора фермента, оставляют в течении 10 мин., добавляют 10 мл 0.3 н. раствора трихлоруксусной кислоты, энергично взбалтывают, фильтруют и 5 мл фильтрата отливают в мерную колбу на 25 мл. Сюда же приливают 10 мл 0.5 н. раствора едкого натра и 3 мл фенольного реактива (реактив Фолина-Чиокалтеу разводят предварительно двойным объемом воды) и доводят водой до метки. По истечении нескольких минут окрашенный раствор фотометрируют относительно стандартного раствора, содержащего 0.0008 мэкв тирозина в 5 мл 0.2 н. раствора соляной кислоты (к раствору добавляют 0.5% формальдегида в качестве антисептика). Для исключения возможных ошибок ставят холостой опыт. Единицы пепсина выражают в миллиэквивалентах тирозина (в пределах от 6.10-4 до 11.10-4): единицы пепсина = миллиэквиваленты тирозина х1.47.
Определение протеолитических ферментов по Муру и Штейну. Метод основан на том, что содержащие a-аминогруппу аминокислоты дают с нингидридом окрашенную производную - дикетогидриндилидендикетогидриндамин, а также альдегид-аминокислоты и углекислоту.
Окрашенный продукт обладает характерным максимумом поглощения при 570 мm, а интенсивность окраски зависит от количества аминокислоты. С пролином и оксипиролином реакция протекает в ином направлении, а максимум поглощения получаемого при этом окрашенного продукта расположен при 440 мm.
Инкубацию фермента удобно проводить в мерных колбочках по 5 мл, в которых смешивают и уравновешивают при постоянной температуре на водяной бане все компоненты за исключением фермента. После этого добавляют фермент, раствор смешивают и берут пробы для исследования. Для кинетических измерений концентрацию фермента желательно подобрать таким образом, чтобы аликвотные части можно было бы брать каждые 5 минут.
Аликвотные части пробы, взятые сразу же после добавления фермента, а затем через соответствующие интервалы, добавляют в фотометрические пробирки, содержащие 2 мл реактива нингидрида - это тотчас обрывает реакцию. После этого смесь 20 мин. нагревают на водяной бане; полученная окраска устойчива минимум 24 часа. Средняя ошибка при анализе повторных проб составляет ±2%. После разбавления кипяченой смеси пробы и нингидрида 10 мл смеси, состоящей из равных объемов воды и н.пропанола, интенсивность окраски определяют на спектрофотометре Колемана при 570 мm.
Для калибровки пользуются постоянными количествами стандартных аминокислот, содержащих реакционные компоненты. При помощи калибровочных кривых, построенных отдельно для каждой исследуемой аминокислоты, измерения интенсивности окраски (по отношению к контрольной колбе) пересчитывают в миллимоли. При делении этих величин на миллимоли субстрата, использованного в процессе реакции, можно найти процент гидролиза.
Реферат опубликован: 1/06/2005 (13927 прочтено)