Страница: 4/5
По своему механизму все фотосенсибилизируемые реакции разделяют на два типа в зависимости от того, каким способом энергия светового возбуждения передается от сенсибилизатора к биологическому субстрату. В реакциях типа I сенсибилизатор в возбужденном триплетном состоянии участвует в переносе электрона или атома водорода, в результате чего получаются реакционноспособные радикалы биологического субстрата, вступающие в дальнейшие химические реакции с кислородом или с другими молекулами. В реакциях типа II происходит перенос энергии от сенсибилизатора к молекуле кислорода с переводом ее в электронно-возбужденное синглетное состояние 1О2; образовавшийся синглетный кислород затем окисляет биологический субстрат. Тип фотосенсибилизируемой реакции зависит в первую очередь от природы сенсибилизатора и субстрата, в частности от их способности вступать в окислительно-восстановительные реакции.
Действие света в присутствии сенсибилизаторов направлено на те компоненты клетки, в которых сенсибилизатор локализуется. За исключением небольшого ряда веществ, которые хорошо проникают в ядро клетки и сенсибилизируют фотоповреждение наследственного аппарата, большинство сенсибилизаторов преимущественно взаимодействует с мембранными системами клетки и обуславливает их фотоповреждение, что приводит к нарушению гомеостаза клетки, ее энергообеспечения и других мембраносвязанных функций.
Биологические мембраны являются главной мишенью повреждающего действия света в присутствии, по крайней мере, порфириновых сенсибилизаторов. Следовательно, для того чтобы ответить на вопрос, каковы механизмы сенсибилизированного повреждения клеток, в первую очередь нужно рассмотреть фотоповреждения мембран. К сожалению, на изолированных мембранах выполнены лишь единичные работы, причем все исследования проведены на эритроцитарной мембране. Однако как было показано, эта мембрана является удобной и адекватной моделью биологических мембран вообще, поэтому можно думать, что закономерности, установленные на ней, будут справедливы и для других мембран.
Поскольку главными компонентами биологических мембран являются липиды и белки, характер фотосенсибилизируемых процессов в мембране определяется в значительной мере чувствительностью белков и липидов к сенсибилизированным фотоповреждениям.
В липидах большую чувствительность к сенсибилизированному фотоокислению проявляют ненасыщенные жирные кислоты, насыщенные же жирнокислотные цепи практически не подвергаются фотоокислению. Легко фотоокисляется также другой вид липидов - холестерин. Процессы фотоокисления ненасыщенных жирных кислот фосфолипидов, а также холестерина сходны с процессами, протекающими при перекисном окислении липидов.
Сенсибилизированные фотоповреждения липидов и белков мембран приводят к изменениям их важнейших функциональных характеристик: снижению барьерных свойств, подавлению активностей мембраносвязанных ферментов и транспортных систем.
Большое значение в фотоиндуцируемом снижении барьерных свойств мембран придается фотоповреждениям липидов. Фотосенсибилизируемое ингибирование ряда мембранных ферментативных активностей связано с прямым фотоповреждением белков. Нарушения барьерных свойств мембран и ее транспортных свойств приводят к изменению внутриклеточного состава и к неизбежной гибели клетки. Можно представить следующую схему фотосенсибилизированных процессов в мембране (3):
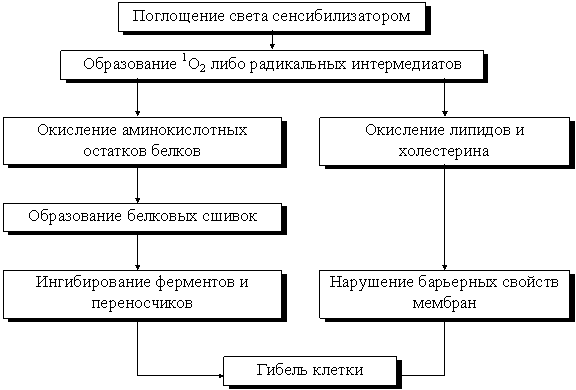
Фотосенсибилизированные повреждения мембран можно ослабить или усилить рядом веществ. Хорошими тушителями синглетного кислорода являются b-каротин, a-токоферол, азид натрия а др. Известны также соединения, которые увеличивают фотосенсибилизируемые повреждения. Это прооксиданты - вещества, которые усиливают разветвления цепи окисления, вступая в окислительно-восстановительные реакции с липидными перекисями (3).
Фотохимический механизм повреждающего действия
света на фоторецепторы сетчатки глаза.
В некоторых экстремальных ситуациях видимый свет, будучи слишком ярким или длительно действующим, способен оказать повреждающее действие на глаз, особенно больной или предрасположенный к заболеванию. Для человека эта проблема становится все более актуальной в связи с появлением техники, имеющей мощное и концентрированное световое излучение. Для поиска средств, предотвращающих отрицательное действие видимого света на глаз человека, необходимо знать, какие ткани глаза при этом повреждаются, и вскрыть механизмы процессов вызывающих фотоповреждение.
Работы последнего времени показали, что фотоповреждение может приводить к дегенерации клеточных элементов сетчатки, в первую очередь ее рецепторов и клеток пигментного эпителия. Наиболее подвержены повреждению фоторецепторные клетки сетчатки - палочки и колбочки. Так, через сутки после достаточно длительного действия на крыс света умеренной интенсивности наблюдали выраженные патологические изменения фоторецепторов. С помощью электронного микроскопа было обнаружено набухание и удлинение их наружных сегментов, дезориентация фоторецепторных дисков. Аномальное удлинение наружных сегментов является, вероятно, следствием нарушения фагоцитоза обломков наружных сегментов, содержащие “старые” диски.
Таким образом, наиболее выраженным является нарушение клеточных контактов между наружными сегментами фоторецепторов и клетками пигментного эпителия. В результате этого в щели между сетчаткой и пигментным эпителием накапливаются непереваренные обломки наружных сегментов. Это может приводить к развитию аутоиммунного воспалительного процесса, например симпатического воспаления глаза.
Через неделю после длительного действия на животное яркого света слой фоторецепторов, а также клеток пигментного эпителия в макулярной области полностью отсутствует.
Эти морфологические данные подтверждены результатами электрофизиологических исследований. Как известно, суммарная электрическая реакция сетчатки - электроретинограмма (ЭРГ) достаточно адекватно отражает функциональное состояние ее клеточных элементов - рецепторных, нервных, глиальных. Существенный вклад в генерацию “а”-волны ЭРГ вносят фоторецепторы. Основной результат электрофизиологических опытов сводится к следующему: при действии на глаз кроликов и крыс повреждающих доз видимого света наиболее чувствительной к фотоповреждению является именно “а”-волна ЭРГ (4).
Интересно проследить динамику раннего рецепторного потенциала (РРП), который, как известно, адекватно отражает содержание в сетчатке необесцвеченных зрительных пигментов. У животных со смешанной сетчаткой основной вклад в генерацию РРП вносят колбочки. Оказалось, что после фотоповреждения РРП восстанавливается быстрее, чем в палочках накапливается зрительный пигмент. Отсюда следует, что более подвержены повреждению ответственные за сумеречное зрение палочки (4).
Развитие патологического процесса идет по механизму фотосенсибилизированного свободнорадикального окисления, так как в зрительных клетках присутствуют три необходимых для этого ингредиента: активно поглощающие свет окрашенные соединения, кислород и субстраты окисления - белки и липиды.
Коротко остановимся на каждом из трех факторов. Начнем со свободного кислорода. Из широкой сети капилляров мощной сосудистой оболочки глаза он диффундирует через однослойный пигментный эпителий к фоторецепторам сетчатки. В ткани сетчатки, как известно парциальное давление кислорода весьма велико, причем около 60% его потребляется именно фоторецепторными клетками. На свету концентрация свободного кислорода в наружном сегменте фоторецепторной клетки возрастает. Следовательно, для развития окислительного процесса в рецепторных клетках нет недостатка в свободном кислороде.
Следующий необходимый для развития процесса фотоокисления фактор - это окрашенные, поглощающие свет вещества: родопсин в палочках, йодопсин в колбочках и продукты их фотолиза. Все зрительные пигменты являются хромогликопротеинами, в которых в роли окрашивающего белок хромофора выступает альдегид витамина А - ретиналь. Необесцвеченнная молекула родопсина поглощает свет в зеленой области спектра (500 нм), а при поглощении кванта света она быстро обесцвечивается, при этом максимум поглощения смещается в синюю область. Именно ретиналь в обесцвеченном зрительном пигменте выступает в качестве фотосенсибилизатора процессов окисления белков и липидов фоторецепторной мембраны наружного сегмента зрительной клетки.
Третьим необходимым компонентом для развития в клетке патологического фотоокислительного процесса являются субстраты окисления, т.е. вещества, которые претерпевают окислительное разрушение. Как известно, фоторецепторная мембрана состоит из липидов (50% сухого веса) и белка (40% сухого веса). Причем она крайне гомогенна по белковому составу - около 60-80% всех белков составляет родопсин. Это - водонерастворимый трансмембранный белок, содержащий шесть свободных тиоловых (SH) групп. В то же время хорошо известно, что в биологической мембране в первую очередь окисляются тиоловые группы белков. Кроме того, в фоторецепторной мембране имеется еще один легко подверженный окислению субстрат - это ненасыщенные жирные кислоты, скорость окисления которых возрастает с увеличением в них числа двойных связей. Для нормального функционирования зрительной клетки крайне важно поддержание малой вязкости фоторецепторной мембраны, которая обеспечивается именно высоким содержанием полиненасыщенных жирных кислот. В фосфолипидах фоторецепторной мембраны преобладает докозагексаеновая жирная кислота, содержащая шесть двойных связей.
Реферат опубликован: 7/04/2005 (24052 прочтено)