Страница: 1/8
ОГЛАВЛЕНИЕ
СПИСОК СОКРАЩЕНИЙ..............................................................................................3
ВВЕДЕНИЕ.............................................................................................................................4
ОБЗОР ЛИТЕРАТУРЫ......................................................................................................5
СТРОЕНИЕ ИММУНОГЛОБУЛИНОВ...................................................................................5
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ
ГЕНОВ L-ЦЕПЕЙ ИГ У МЛЕКОПИТАЮЩИХ....................................................................7
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ
ГЕНОВ L-ЦЕПЕЙ ИГ У НИЗШИХ ПОЗВОНОЧНЫХ......................................................11
ЭВОЛЮЦИЯ ГЕНОВ L-ЦЕПЕЙ ИГ.......................................................................................19
МАТЕРИАЛЫ И МЕТОДЫ...........................................................................................24
ЭЛЕКТРОФОРЕЗ ДНК............................................................................................................24
ВЫДЕЛЕНИЕ ДНК ИЗ ГЕЛЯ.................................................................................................24
ПОЛУЧЕНИЕ РАДИОАКТИВНЫХ ЗОНДОВ......................................................................25
ВЫДЕЛЕНИЕ ПЛАЗМИДНОЙ ДНК......................................................................................26
ОЧИСТКА ПЛАЗМИДНОЙ ДНК МЕТОДОМ РАВНОВЕСНОГО
ЦЕНТРИФУГИРОВАНИЯ В ГРАДИЕНТЕ ПЛОТНОСТИ CsCl........................................26
ТРАНСФОРМАЦИЯ ПРОКАРИОТИЧЕСКИХ КЛЕТОК...................................................27
ВЫДЕЛЕНИЕ ЛЕЙКОЦИТОВ ИЗ КРОВИ РЫБ.................................................................28
ВЫДЕЛЕНИЕ СУММАРНОЙ РНК ИЗ ЛЕЙКОЦИТОВ.....................................................28
ВЫДЕЛЕНИЕ ВЫСОКОМОЛЕКУЛЯРНОЙ ДНК
ИЗ ЭУКАРИОТИЧЕСКИХ КЛЕТОК.....................................................................................28
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК......................................................................29
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ...............................................................................33
СУБКЛОНИРОВАНИЕ ДНК...................................................................................................34
СКРИНИНГ БИБЛИОТЕКИ кДНК........................................................................................34
ОПРЕДЕЛЕНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК.............................35
САУЗЕРН БЛОТ ГИБРИДИЗАЦИЯ.....................................................................................36
РЕЗУЛЬТАТЫ.....................................................................................................................38
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК И ЕЕ СКРИНИНГ.....................................38
ОПРЕДЕЛЕНИЕ ПЕРВИЧНОЙ СТРУКТУРЫ......................................................................40
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ..............................................40
АНАЛИЗ ГЕНОМНОЙ ОРГАНИЗАЦИИ..............................................................................41
ОБСУЖДЕНИЕ....................................................................................................................46
ВЫВОДЫ...............................................................................................................................48
СПИСОК ЛИТЕРАТУРЫ................................................................................................49
СПИСОК СОКРАЩЕНИЙ
ИГ иммуноглобулины
пн пар нуклеотидов
тпн тысяч пар нуклеотидов
е.а. единица активности
ИПТГ изопропилтиогалактозид
ТЕМЕД N,N,N,'N'-тетраметил-этилен-диамин
трис трис(гидроксиметил)аминометан
дНTФ дезоксиаденозинтрифосфат
ддНТФ дидезоксинеклеотидтрифосфат
дНТФ дезоксинуклеотидтрифосфат
рATФ рибоаденозинтрифосфат
ДТТ дитиотрейтол
Na2ЭДТА этилендиаминтетраацетат натрия
SDS додецилсульфат натрия
X-Gal 5-бромо-4-хлоро-3-индолил-бета-D-галактозид
ВВЕДЕНИЕ
В основе функционирования гуморального иммунитета у млекопитающих лежит сложный комплекс молекулярно-генетических и физиологических механизмов, обеспечивающих реорганизацию, мутагенез и клональную экспрессию генов иммуноглобулинов. Возникновение этой подсистемы иммунитета связывают с появлением хрящевых рыб.
На ранних этапах эволюции гены ИГ были организованы в виде повторяющихся кластеров V-J-C генных сегментов. При подобной организации разнообразие продуцируемых антител основывалось, прежде всего, на количестве имевшихся в геноме кластеров, каждый из которых кодировал один вариант полипептидных субъединиц ИГ. В ходе дальнейшей эволюции произошел переход от кластерной к сегментарной организации, обеспечивающей дополнительный источник разнообразия за счет комбинативной рекомбинации генных сегментов. Предполагается, что этот переход также имел важное значение для регуляции экспрессии генов и осуществления механизмов клонального отбора антителопродуцирующих клеток в ходе иммунного ответа.
Настоящая работа представляет собой часть проекта, направленного на изучение эволюции механизмов изотипического исключения генов легких цепей ИГ у низших позвоночных. Целями работы являлись изучение структуры и организации генов легких цепей ИГ стерляди (Acipenser ruthenus), представителя филогенетически древней группы костно-хрящевых рыб.
ОБЗОР ЛИТЕРАТУРЫ
СТРОЕНИЕ ИММУНОГЛОБУЛИНОВ
Иммуноглобулины выполняют в организме позвоночных функцию гуморальных антител и антиген-связывающих рецепторов В-лимфоцитов. Особенностью этого класса белков является огромное разнообразие, позволяющее им вступать во взаимодействие с фактически любыми биологическими макромолекулами.
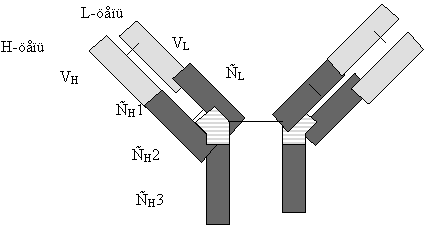
Типичная молекула ИГ состоит из двух идентичных тяжелых (H) и двух идентичных легких (L) полипептидных цепей. H- и L-цепи построены из нескольких доменов, каждый из которых состоит примерно из 110 аминокислотных остатков. У млекопитающих существует пять классов H-цепей: g, m, a, d, и e. Каждая H-цепь построена из одного N-концевого и нескольких (трех или четырех) C-концевых доменов. N-концевые домены различаются в разных молекулах и называются вариабельными (V) доменами. C-концевые домены имеют одинаковую структуру у молекул одного класса и называются константными (С) доменами (рис. 1) (Пол, 1987).
Среди L-цепей млекопитающих различают два типа: лямбда и каппа. Каждая L-цепь состоит из одного вариабельного и одного константного доменов. В индивидуальной молекуле ИГ присутствует только один тип L-цепи (Roitt et al., 1993).
Гигантское разнообразие ИГ обеспечивается прежде всего тем, что V и C области кодируются разными генами (генными сегментами), физически разнесенными в зародышевой ДНК.
В формировании вариабельных доменов H-цепей участвуют три генных сегмента:
вариабельный (V), D-сегмент (от англ. diversity- разнообразие) и J-сегмент (от
англ. joining- соединяющий). С-области Н-цепей разных классов кодируются отдельными
генами (Roitt et al., 1993).
 |
|||
 |
|||
Рис. 1. Схема строения молекулы иммуноглобулина.
Типичная молекула иммуноглобулина содержит две идентичные легкие (L) и две идентичные тяжелые (H) цепи, связанные между собой ковалентно дисульфидными связями. Каждая L-цепь состоит из одного вариабельного (VL) и одного константного (СL) домена. Н-цепь состоит из одного VH и нескольких CН доменов.
В формировании зрелого гена L-цепи участвуют три генных сегмента: V-сегмент и J-сегмент кодируют V-домен, C-сегмент кодирует константный домен. На поздних стадиях развития В-лимфоцита генные сегменты, кодирующие вариабельные домены, объединяются в различных сочетаниях, образуя матрицу для экспрессии L- и H-цепей (Seidman et al., 1979; Durdik et al., 1984).
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ ГЕНОВ L-ЦЕПЕЙ ИГ У МЛЕКОПИТАЮЩИХ
Различные виды млекопитающих имеют сходную схему организации и экспрессии генов ИГ. Одним из наиболее изученных в этом отношении видов является человек. Лямбда и каппа цепи ИГ у человека кодируются генами, расположеными в разных локусах и на разных хромосомах. Каппа локус состоит из большого количества Vk генных сегментов, собраных в группы, пяти Jk и одного Сk сегмента.. Такой тип организации называется сегментарным (рис. 2). Лямбда локус состоит из группы Vl сегментов, точное количество которых неизвестно, и семи пар Jl-Cl сегментов. Три из них (JCl4 - JCl6) являются псевдогенами (Hieter et al., 1981; Roitt et al., 1993).
В процессе развития В-лимфоцита в кроветворных органах один из V сегментов соединяется с одним из J сегментов посредством сайтспецифической рекомбинации (Schatz et al., 1992). В этот процесс вовлекаются специальные олигомерные последовательности: гепта- и нонамеры, фланкирующие с 3' стороны V сегмент и с 5' стороны J сегмент (рис. 3) (Aguilera et al., 1987; Hesse et al., 1989). Отличительной особенностью лямбда и каппа локусов является конфигурация промежутков между гепта- и нонамерами, называемых спейсерами. С V и J генными сегментами лямбда типа ассоциированы 23 пн и 12 пн спейсеры соответственно, а с V и J сегментами каппа типа - 12 пн и 23 пн спейсеры (Durdik et al., 1984; Stavnezer et al., 1985).
|
||||||
| ||||||